№1, 2005 г.
2004 год
НОБЕЛЕВСКАЯ
ПРЕМИЯ
по физиологии и медицине
Р.Эксел и Л.Бак
2004 год
НОБЕЛЕВСКАЯ
ПРЕМИЯ
по физиологии и медицине
Р.Эксел и Л.Бак

Р. Эксел |

Л. Бак |
4 октября 2004 г. Нобелевский комитет по физиологии и медицине в Каролинском институте (Стокгольм) объявил о присуждении премии двум американским исследователям Р.Экселу и Л.Бак “за открытие обонятельных рецепторных белков и организации обонятельной системы”.
Ричард Эксел (Richard Axel) родился в 1946 г. в Нью-Йорке. В 1967 г. окончил Колумбийский университет, докторскую степень получил в 1970 г. в Университете Джона Хопкинса (Балтимор), а звание профессора - в 1978 г. в Колумбийском университете (Нью-Йорк), где и работает до настоящего времени.
Линда Бак (Linda B.Buck) родилась в Сиэтле в 1947 г. Окончила Вашингтонский университет в своем родном городе в 1975 г. по специальности физиолог, микробиолог. Докторскую диссертацию защитила в 1980 г. в Техасском университете (Даллас). С 1984 по 1991 г. работала в Колумбийском университете (Нью-Йорк), затем была ассистентом профессора (с 2001 г. профессором) отделения нейробиологии Гарвардской медицинской школы (Бостон). С 2003 г. - профессор отделения физиологии и биофизики Вашингтонского университета в Сиэтле.
Чтобы понять причины столь высокой оценки работы двух американских ученых, полезно кратко ознакомиться с ретроспективой исследований в этой области. С древнейших времен людей интересовало, как устроены и как работают органы чувств человека и животных. Еще почти 2 тыс. лет назад, в середине I в. до н.э., древнеримский философ и поэт Тит Лукреций Кар, ученик Эпикура, составил энциклопедию естественнонаучных представлений своего времени. Сочинение Лукреция “О природе вещей”, написанное в поэтической форме (что прославило автора вдвойне), состоит из шести книг, одна из которых (книга IV) почти полностью посвящена механизмам работы органов чувств. Примерно 80% объема книги отведены зрению и слуху, и всего 20% химическим чувствам - вкусу и обонянию. Любопытно, что и в наше время, т.е. спустя 2 тыс. лет, примерно такое же распределение материала продолжает сохраняться в научных обзорах и монографиях, посвященных органам чувств. Вероятно, таково соотношение значимости отдельных органов чувств в жизни человека, и, соответственно, приоритетов в экспериментальных исследованиях разных видов рецепции. Если бы книги про органы чувств писали не люди, а представители животного мира, то в центре внимания в подавляющем большинстве случаев, скорее всего, оказалось бы обоняние.
В XX в. наметился заметный контраст между успехами в изучении зрительной системы, где активно использовался непрерывно расширяющийся арсенал современных методик, и более медленным прогрессом в исследованиях обонятельной рецепции, которые проводились усилиями сравнительно малочисленной группы энтузиастов в разных странах. Успехи активной работы по изучению зрения были дважды (1967 и 1981) отмечены Нобелевскими премиями. После этих наград, полученных коллегами по сенсорному цеху, работы по изучению обоняния оживились. Число лабораторий, вовлеченных в исследование обонятельной рецепции, число публикаций и объем знаний в этой области стали быстро возрастать. Уже в начале 70-х годов были отброшены некогда популярные полуфантастические гипотезы, приписывающие обонятельным клеткам уникальные механизмы рецепции, вроде усиления на основе сегнетоэлектрических свойств или излучения электромагнитных волн. Большинство исследователей по аналогии с другими, лучше изученными реакциями клеток на химические сигналы (например, рецепцией гормонов или синаптических нейромедиаторов) предполагало, что в мембране обонятельных клеток присутствуют специальные белки-рецепторы, а их взаимодействие с молекулами пахучих веществ приводит к изменению электрического потенциала клеточной мембраны. Такое преобразование первичного сенсорного сигнала в стандартный ответ рецепторных клеток (сдвиг трансмембранного потенциала) назвали трансдукцией.
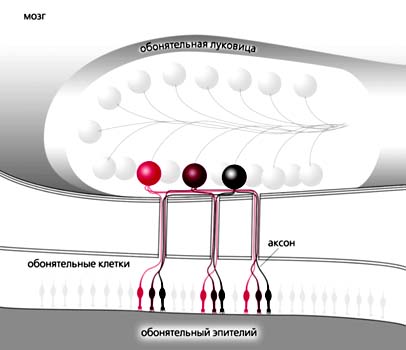
К концу XX в. особенности морфологии и физиологии обонятельной системы были изучены достаточно хорошо, как у животных, так и у человека. В составе обонятельной системы различают периферический отдел, проводящие пути и обонятельные центры мозга. У позвоночных животных периферический отдел представлен обонятельным эпителием, располагающимся в полости носа; у человека - в верхних отделах средней носовой раковины, верхней носовой раковине и верхней части перегородки носа. Непосредственно с молекулами пахучих веществ взаимодействуют биполярные обонятельные рецепторные клетки (их также называют обонятельными рецепторными нейронами, поскольку они во многом похожи на настоящие нервные клетки). От вершины обонятельной клетки отходит периферический отросток (дендрит), отличающийся, однако, от дендритов нервных клеток. У обонятельной клетки он заканчивается вздутием (булавой), которое несет пучок длинных тонких ресничек, напоминающих жгутики простейших. Булавы и реснички погружены в слой слизи, покрывающий обонятельную выстилку. Как и у большинства нервных клеток, у обонятельной рецепторной клетки имеется также длинный тонкий центральный отросток, аксон, по которому возбужденная клетка посылает в обонятельный центр мозга (обонятельную луковицу) серию нервных импульсов. На поверхности обонятельной луковицы расположены особые синаптические структуры, обонятельные клубочки, в каждом из которых происходит передача сигналов от десятков тысяч аксонов рецепторных клеток к дендритам нескольких десятков митральных (и некоторых других) нейронов обонятельной луковицы. Обонятельная система беспозвоночных построена по сходному принципу: там тоже аксоны большого числа обонятельных рецепторных клеток объединяются в клубочки в обонятельных центрах.
Функциональная схема обонятельной системы. Снизу вверх. На поверхности обонятельного эпителия молекулы пахучего вещества взаимодействуют с миллионами обонятельных рецепторных клеток. Каждая клетка несет на ресничках молекулы какого-то одного из тысячи возможных рецепторных белков, который определяет характер ее чувствительности к разным запахам. Длинные отростки рецепторных клеток (аксоны) подходят к мозговому центру обоняния - обонятельной луковице. На ее поверхности аксоны перераспределяются так, чтобы сигналы от клеток с одинаковыми рецепторными белками собирались в одном клубочке. Нейронные сети обонятельной луковицы анализируют пространственную картину распределения возбужденных клубочков и передают информацию о запахе в отделы мозга, управляющие пищевым, половым, социальным и другими типами поведения животного. (По: R.Axel, 1995.)Особенность обонятельных рецепторных клеток - ограниченная продолжительность их жизни. У позвоночных они постоянно отмирают и заменяются новыми, которые дифференцируются в результате митотического деления базальных (стволовых) клеток обонятельного эпителия. Таким образом, связи периферического отдела обонятельной системы с мозгом - динамические: старые синаптические контакты постоянно разрушаются, а подрастающие аксоны рецепторных клеток образуют новые. В ходе такого непрерывного “обновления” восприятие и узнавание запахов не нарушается
Несмотря на накопленный большой экспериментальный материал, механизм обонятельной рецепции на молекулярном уровне, так же как и принципы кодирования обонятельной информации в нервной системе, позволяющие воспринимать и узнавать огромное количество запахов, оставались во многом загадкой для ученых. Из электрофизиологических опытов было известно, что обонятельные клетки высоко чувствительны к одним пахучим веществам и малочувствительны к другим, причем разные клетки обладают разными “спектрами чувствительности”. Представлялось очевидным, что мозг не мог бы определить качество обонятельного стимула, воспринимаемого рецепторными клетками, если бы сигналы от рецепторных клеток всех типов равномерно смешивались и усреднялись в клубочках. Уже в 1957 г. У.Легро-Кларк предложил гипотезу, согласно которой связь обонятельных рецепторных клеток с обонятельной луковицей мозга должна быть упорядочена так, чтобы клетки с одинаковыми “спектрами чувствительности” (т.е. несущие одинаковые рецепторные белки) посылали свои аксоны в один и тот же клубочек обонятельной луковицы [1].
Если отдельный обонятельный клубочек рассматривать как функциональную единицу, то каждому обонятельному стимулу соответствует особая картина (пространственный паттерн) из сильно или слабо возбужденных клубочков. Такое кодирование обеспечивает возможность различения и узнавания огромного числа запахов. У млекопитающих с хорошо развитым обонянием в луковице имеется порядка 1000 клубочков. Даже если у клубочка есть только два состояния (возбужден и не возбужден), общее число различных картин, включая нулевую (отсутствие запаха), составит 21000, или примерно 10300, т.е. будет невообразимо огромным. Гипотеза Легро-Кларка получила косвенное подтверждение в опытах французских физиологов П.Маклеода и Ж.Левето. Им удалось с помощью специальных микроэлектродов зарегистрировать слабый электрический ток, который создает возбужденный обонятельный клубочек. Используя эту методику, они впервые описали специфические пространственные картины в слое обонятельных клубочков в ответ на стимуляцию разными запахами [2].
Позднее пространственные картины в луковице, которыми кодируются запахи в обонятельной системе, удалось наблюдать в нескольких лабораториях при изучении метаболической активности клубочков и клеток луковицы с использованием меченой дезоксиглюкозы [3, 4]. С тех пор предложенная Легро-Кларком гипотеза стала общепринятой, хотя, строго говоря, ее справедливость была доказана только в 90-е годы прошлого века.
В 70-80-х годах в значительной степени выяснились основные этапы обонятельной трансдукции. Исследования, проведенные в лаборатории М.А.Островского в Институте химической физики РАН в Москве и затем в лабораториях Д.Лансета (Израиль), Г.Шеперда (США) и ряда других, установили, что в результате взаимодействия обонятельной клетки с пахучим веществом в клетке появляется особое сигнальное вещество (“вторичный мессенджер”). Его молекулы связываются с ионными каналами в клеточной мембране, увеличивая их проницаемость для катионов. Это приводит к деполяризации клеточной мембраны и появлению или усилению импульсной активности в аксоне клетки. Было также показано, что гипотетический обонятельный рецепторный белок действует через так называемый ГТФ-связывающий белок (такие белки, также называемые G-белками, образуют особую группу белков, которые участвуют в качестве посредников во многих процессах клеточной рецепции). В обонятельных клетках позвоночных в качестве вторичного мессенджера обычно работает циклический аденозин-3’-5’-монофосфат (АМФ). Это хорошо известное вещество выполняет роль вторичного мессенджера во многих процессах клеточной рецепции, в частности, в рецепции гормонов. В обонятельных клетках описаны и другие мессенджеры, в том числе ионы Ca2+, которые тоже регулируют проницаемость ионных каналов.
Фактически в цепочке обонятельной трансдукции были надежно идентифицированы все звенья, кроме начального - обонятельных рецепторных белков, несмотря на особое внимание, которое им уделялось. В многочисленных попытках обнаружить их среди мембранных белков обонятельного эпителия особое внимание уделялось способности связывать пахучие вещества с достаточно низкой величиной константы диссоциации (Kd) - ключевой признак рецепторного белка. Значительная часть биохимических и электрофизиологических работ была посвящена поиску связывающих аминокислоты рецепторных белков рыб. В лаборатории Е.Е.Фесенко (в Институте биофизики клетки РАН, 1980, 1983, 1989 гг.) из обонятельной ткани ската и карпа выделили белок с центрами связывания, имеющими очень высокое сродство к соответствующим L-аминокислотам (Kd = 10–10 М). Эти рецепторные белки ассоциированы с ГТФ-связывающим белком. В этой же лаборатории из поверхностного слоя обонятельной выстилки крысы получили мембранный белок gp88 (молекулярная масса 88 кДа), связывающий пахучие вещества (камфору, деканаль), а в мембране ассоциированный с ГТФ-связывающим белком. Моноклональные антитела к gp88 подавляли связывание пахучих веществ и вызываемый ими суммарный электрический ответ обонятельной выстилки. Еще один кандидат в обонятельные рецепторные белки - гликопротеид gp95, выделенный в лаборатории Д.Лансета из обонятельных ресничек нескольких видов животных. Он также удовлетворял большей части биохимических критериев, определяющих рецепторную функцию мембранных белков. Все эти белки были получены в небольших количествах, что затрудняло более глубокий анализ их свойств, а их рецепторная функция не была доказана достаточно убедительно для научного сообщества.
На фоне этих успехов проблема остающихся неуловимыми обонятельных рецепторных белков, особенно при сравнении с детально изученным зрительным белком родопсином, стала настоящим вызовом для “обонятельного” научного сообщества и, вероятно, для молекулярной биологии в целом. Ответом на него оказалась работа, выполненная в лаборатории профессора Колумбийского университета Р.Эксела (США), результаты которой были опубликованы в 1991 г. [5]. Авторы искали решение задачи обходным путем, поставив целью описать свойства рецепторных белков на языке кодирующих их генов.
Известно, что клетки синтезируют необходимые для них белки на основе генетической информации, записанной в ДНК. В соответствии с этой информацией в ходе транскрипции строится матричная РНК (мРНК), которая используется в качестве шаблона при соединении аминокислот в белковую молекулу. Методы молекулярной биологии позволяют обратить этот процесс, так что, выделив из клеток мРНК, с помощью обратной транскрипции удается клонировать соответствующие гены. Но в каждой клетке имеется множество белков и молекул мРНК. Чтобы сократить перебор, авторы использовали три ожидаемых свойства обонятельных рецепторных белков:
В своей первой публикации авторы сообщили об открытии нового семейства генов у крысы, содержащего не менее (скорее всего, более) 100 членов, которые (как осторожно указано в заголовке) “возможно, кодируют обонятельные рецепторы”.
- число типов обонятельных белков очень велико, и, соответственно, семейство обонятельных генов должно быть достаточно многочисленным;
- участие G-белка в возбуждении обонятельных клеток означает, что искомые белки принадлежат к известной группе белковых рецепторов, сопряженных с G-белками. Все такие белки имеют характерные детали молекулярного строения (например, семь петель полипептидной цепи, которые пересекают клеточную мембрану). Соответственно, гены, кодирующие эти белки, имеют характерные участки, по которым их можно опознать;
- экспрессия обонятельных рецепторных белков должна быть ограничена обонятельным эпителием.
Работа получила широкий резонанс (ее называли примером “триумфа молекулярной биологии”). Она как бы открыла створки шлюза - вслед за ней в течение короткого промежутка времени из разных лабораторий последовали десятки публикаций, описывающих свойства обонятельных генов у разных видов животных и у человека, локализацию их в хромосомах и т.д. В ряде работ сообщались уточненные оценки репертуара обонятельных генов для крысы и мыши: примерно 500-1000 (против первоначальной оценки 100-200).
У человека это семейство насчитывает более 500 генов, однако большая их часть (до 3/4 от общего числа) не функционирует. Это так называемые псевдогены, потерявшие способность управлять синтезом полипептидов. У исследованных животных такие псевдогены не обнаружены. Утрата человеком большей части обонятельных генов и, соответственно, рецепторных белков может быть связана с тем, что эволюция Homo sapiens шла в направлении ослабления роли обоняния по сравнению со зрением и слухом. Если в популяции людей отдельные индивидуумы имеют разные псевдогены, это должно сказаться на их обонятельных ощущениях, что, возможно, объясняет удивительный разброс между людьми в оценках интенсивности и качества предъявляемых запахов.
Вслед за работой Бак и Эксела семейства обонятельных генов были описаны для собаки, свиньи, опоссума, курицы, лягушки, протея и нескольких видов рыб (у них всего около 100 генов). Помимо обонятельного эпителия, обонятельные гены были также обнаружены в семенниках млекопитающих (что, вероятно, связано с рецепцией сперматозоидов, отыскивающих яйцеклетку по химическим сигналам). Был исследован вомероназальный орган млекопитающих, в котором обнаружено два независимых семейства обонятельных генов, оба - отличные от генов основного обонятельного эпителия [6]. Среди насекомых семейство обонятельных генов исследовано у классического объекта генетиков - дрозофилы [7, 8].
Таким образом, возникло новое направление исследований, родоначальники которого, Ричард Эксел и Линда Бак, удостоены высшей научной награды. Эта оценка фактически означает что-то вроде волшебного превращения обонятельной системы из Золушки в равноправную принцессу среди других сенсорных систем.
Казалось бы, клонирование обонятельных генов и расшифровка их структуры фактически решали задачу получения обонятельных рецепторных белков и исследования их свойств. В лабораториях для этих целей используют хорошо разработанные методы генной инженерии для экспрессии нужных генов в удобной для манипуляций посторонней клетке-хозяине и “выращивания” в ней интересующего белка. Однако печатных сообщений о таких работах с обонятельными рецепторными белками появилось на удивление мало. В них отмечались неожиданные проблемы, с которыми столкнулись экспериментаторы и которые полностью преодолеть не удалось. В основном, они сводились к тому, что при экспрессии обонятельных генов не было никаких признаков появления соответствующих рецепторных белков в клеточной мембране. Если учесть, что в современной практике научных публикаций сообщать о неудачных опытах обычно не принято, можно предположить, что фактически таких неудачных попыток было гораздо больше. Таким образом, проблема обонятельных рецепторных белков опять повисла в воздухе.
Тем временем в лаборатории Р.Эксела и независимо в новой лаборатории, возглавляемой Линдой Бак, был получен еще один сенсационный результат. Используя изощренную технику манипуляций с генами, экспериментаторам удалось связать экспрессию трех обонятельных генов в обонятельных клетках эпителия с тремя разными цветным маркерами. Это позволило проследить организацию контактов аксонов обонятельных рецепторных клеток с клубочками обонятельной луковицы. Полученные картины однозначно свидетельствовали о том, что терминали аксонов рецепторных клеток, в которых имеются одинаковые обонятельные гены, собираются в одном и том же клубочке [9].
В этот же период появилось весьма наглядное подтверждение гипотезы о пространственном кодировании запахов в клубочковом слое обонятельной луковицы. Несколькими лабораториями в США и Германии были успешно применены оптические методы с использованием флуоресцентных красителей, позволяющих придавать специальную окраску возбужденным клубочкам. Были описаны специфические пространственные паттерны в слое обонятельных клубочков, вызываемые обонятельными стимулами, у млекопитающих [10], рыб [11] и насекомых [12]. Особенно эффектно выглядели опыты, проведенные в лаборатории К.Менцеля в Берлинском университете [13]. В эксперименте использовали пчел, у которых число клубочков невелико и каждый из них имеет определенное положение в надглоточном ганглии пчелы, одинаковое у всех особей, так что клубочки легко “узнаются в лицо”. Пчелу обдували воздухом с добавлением разных пахучих веществ. Глядя только на картину расположения возбужденных клубочков, экспериментатор мог сразу же определить, какой именно запах использовался для стимуляции.
Но как же быть со столь упорно ускользавшими обонятельными рецепторными белками? Ситуация в некоторых отношениях еще более усложнилась. После доказательства пространственного кодирования запахов в клубочках выяснилось, что правильная ориентация растущих аксонов вообще невозможна без участия обонятельных генов, которые экспрессируются не только в теле обонятельной клетки, но и в ее аксоне. Сразу же было отмечено, что эти наблюдения имеют прямое отношение к фундаментальным проблемам онтогенеза, в ходе которого направленные перемещения клеток и рост нервных волокон управляются адресными химическими сигналами окружающих тканей.
В таком контексте уместен вопрос: что же такое обонятельные гены и рецепторные белки - специфические детали механизма обоняния или представители другого, пока не изученного, обширного семейства генов и молекулярных рецепторов, обеспечивающих управляемые адресным кодом процессы? В этой связи У.Дрейер из Калифорнийского технологического института отмечает, что локализация так называемых обонятельных генов в действительности отнюдь не ограничена обонятельными клетками, поскольку при изучении существующих генетических баз данных оказалось, что гены из этого семейства фактически обнаруживались в большом числе различных органов и тканей, не имеющих отношения к обонятельной системе.
В связи со всем этим возникают новые вопросы, в частности, можно ли приписывать одному и тому же обонятельному рецепторному белку обе функции - и рецепцию попадающих извне низкомолекулярных пахучих веществ, и рецепцию химических сигналов, которые служат индивидуальными опознавательными метками тканей и клеток, и которые, очевидно, имеют совсем иную химическую природу, чем обычные обонятельные стимулы? Или неуловимые обонятельные рецепторные белки снова ускользают из рук?
© Минор А.В.,
кандидат биологических наук
Институт проблем экологии и эволюции им.А.Н.Северцова РАНЛитература
1. Le Gros Clark W.E. // Proc. Roy. Soc. 1957. V.146B. P.299-318.
2. Leveteau J., Mac Leod P. // J. Physiol. 1969. V.61. P.5-16.
3. Lancet D. et al. // PNAS. 1982. V.79. P.670-674.
4. Benson T.E. et al. // Brain Res. 1985. V.339. P.67-78.
5. Buck L., Axel R. // Cell. 1991. V.65. P.175-187.
6. Mombaerts P. // Science. 2001. V.286. P.707.
7. Gao Q., Chess A. // Genomics. 1999. V.60. P.31-39.
8. Clyne P. et al. // Neuron. 1999. V.22. P.327-338.
9. Mombaerts P. et al. // Cell. 1996. V.87. P.675-686.
10. Rubin B.D., Katz L.C. // Neuron. 1999. V.23. P.499-511.
11. Friedrich R.W., Korsching S.I. // Neuron. 1997. V.18. P.737-752.
12. Korsching S.I. // Cell. 2001. V.58. P.520-530.
13. Jorges J., Kuettner J., Galizia A., Menzel C. // Nature. 1997. V.387. P.85-88.