№2, 2004 г.
© Ковальзон В.М.
МЕЛАТОНИН - БЕЗ ЧУДЕС
В.М. Ковальзон
Владимир Матвеевич Ковальзон - доктор биологических наук, ведущий научный сотрудник
Института проблем экологии и эволюции им. А.Н.Северцова РАНМелатонин - одно из эволюционно древнейших биохимических веществ - был открыт американским дерматологом А.Лернером лишь полвека назад. Это вещество присутствует уже у одноклеточных организмов и у растений, а значит, и в обычной растительной пище, но в ничтожных, “гомеопатических” концентрациях, никак не влияющих на организм млекопитающего. Например, чтобы повысить уровень мелатонина в крови хотя бы до 50 пг/мл (его “вечернее” содержание у человека), нужно съесть за один прием килограммов 200 помидоров или бананов или полтонны отварного риса!
В организме позвоночных животных главный источник мелатонина - эпифиз, или шишковидная (пинеальная) железа. Считают, что эпифиз впервые описал александрийский врач Герофил за 300 лет до н.э., а название свое он получил от Галена (II век н.э.), которому форма железы напомнила сосновую шишку. В XVII в. Р.Декарт приписывал эпифизу роль “седалища души” и связывал его функции со зрением, что весьма интересно в свете современных знаний. Однако на протяжении XVIII-XIX вв. эпифиз рассматривали лишь как рудиментарный придаток мозга. Только в самом конце XIX в. немецкий педиатр О.Хюбнер описал мальчика, отличавшегося преждевременным половым созреванием, у которого при посмертном вскрытии обнаружили опухоль эпифиза. Как теперь очевидно, она препятствовала выработке мелатонина. В начале XX в. невролог О.Марбург предположил, что эпифиз - верхний придаток мозга, выделяет какое-то вещество, угнетающее функции гипоталамуса (важнейшей структуры в глубине мозга, управляющей нижним придатком мозга, гипофизом) и, как следствие - развитие репродуктивной системы. Примерно тогда же установили, что эпифиз содержит субстанцию, вызывающую депигментацию (побледнение) кожи головастиков. Через 40 лет этот факт сыграл решающую роль в открытии мелатонина. Интересно, что первооткрыватель мелатонина Лернер впервые описал и его седативный (успокоительный) эффект при введении человеку.
Мелатонин и эпифиз
У зародыша эпифиз образуется из выпячивания крыши межуточного мозга, из которого берут свое начало и глаза, и гипоталамус. Исторически все эти образования возникли как единое целое - некий механизм, способный реагировать на циклические изменения в световом режиме. У холоднокровных позвоночных и у птиц эпифиз выполняет хорошо известную роль “третьего глаза”, снабжая организм этих животных информацией о суточной и сезонной освещенности. Однако у млекопитающих верхний мозговой придаток, “погребенный” под разросшимися полушариями и мощным черепом, потерял непосредственные афферентные (центростремительные) и эфферентные (центробежные) связи с мозгом и превратился в железу внутренней секреции. Так случилось у всех млекопитающих, за исключением неполнозубых (муравьедов, ленивцев), панцирных (броненосцев) и китообразных (китов, дельфинов), у которых эпифиз попросту исчез.
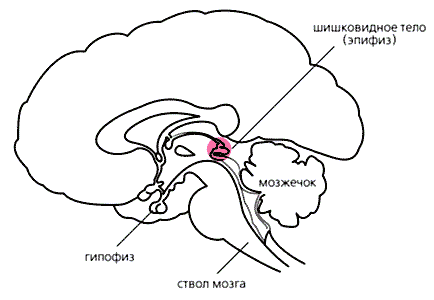
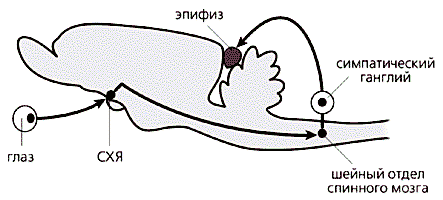
Рис.1. Схема расположения эпифиза (продольный вертикальный срез головного мозга человека). Несмотря на то, что геометрически эпифиз располагается в самом центре мозга, управляется он, как обычный периферический орган, при помощи вегетативной нервной системы (рис.1). Зрительная информация от сетчатки через ответвление зрительного нерва попадает в супрахиазмальные ядра (СХЯ), находящиеся в глубине полушарий над зрительным перекрестом. Затем эти сигналы нисходят вниз (через гипоталамус по проводящим путям вдоль ствола головного мозга), в шейный отдел спинного мозга, откуда по симпатическим нервам через отверстия в черепе проникают обратно в головной мозг и, наконец, достигают эпифиза (рис.2). Ночью, в темноте, когда большинство нейронов супрахиазмальных ядер бездействует, эти нервные окончания выделяют норадреналин, активирующий в клетках эпифиза (пинеалоцитах) синтез ферментов, образующих мелатонин. Эпифиз здорового взрослого человека, имеющий массу немногим более 100 мг, еженощно выделяет в кровь около 30 мкг мелатонина. Яркий свет мгновенно блокирует его синтез, в то время как в постоянной темноте суточный ритм выброса, поддерживаемый периодической активностью СХЯ, сохраняется.
Рис.2. Схема нервной регуляции эпифиза в мозге млекопитающего (продольный вертикальный срез головного мозга крысы). Мелатонин синтезируется из незаменимой (для человека) аминокислоты триптофана, поступающей в организм с пищей. Попав с кровотоком в эпифиз, эта аминокислота превращается в серотонин в два этапа, с участием ферментов триптофангидроксилазы и 5-окситриптофандекарбоксилазы. Затем, также в две стадии, с помощью ферментов N-ацетилтрансферазы (NAT) и оксииндол-О-метилтрансферазы (HIOMT) из серотонина образуется мелатонин (рис.3). Синтезируемое вещество не накапливается в эпифизе, а немедленно выбрасывается в кровяное русло и ликвор (спинномозговую жидкость). Чем ближе к эпифизу, тем выше концентрация мелатонина в биологических жидкостях. Например, ночью в ликворе боковых желудочков его в семь, а в ликворе третьего желудочка - в 20 раз больше, чем в плазме крови, взятой из яремной вены овцы. Однако появление эпифизарного гормона в системной крови отнюдь не результат его простой диффузии из ликвора: ночной подъем мелатонина начинается раньше в крови. Вероятно, эпифиз выбрасывает гормон в виде двух отдельных “порций” (компартментов): одна, в низкой концентрации, поступает в кровь и связывается с периферическими органами и тканями, а другая, в более высоких концентрациях, - в ликвор и связывается с рецепторами мозга.
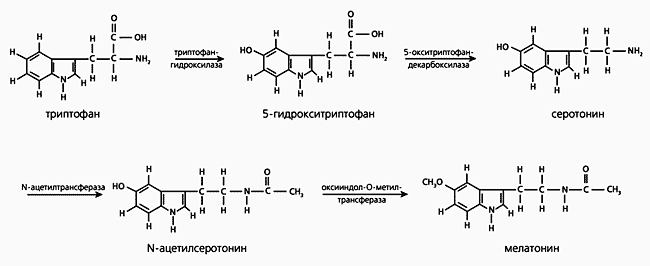
Рис.3. Синтез мелатонина в эпифизе.
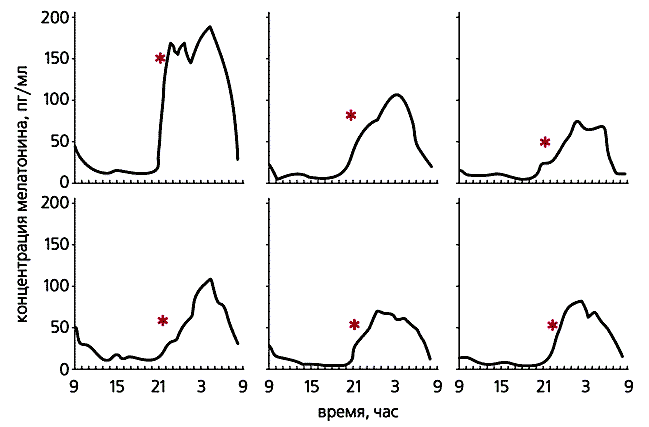
Суточный ход уровня мелатонина в крови (мелатониновая кривая) имеет некоторые сходные черты у всех людей. Так, его концентрация, ничтожная днем (1-3 пг/мл), начинает возрастать часа за два до привычного для данного субъекта времени отхода ко сну (если нет яркого света). После выключения света в спальне концентрация мелатонина быстро увеличивается (до 100-300 пг/мл). В предутренние часы обычно начинается спад, который завершается после пробуждения. Для каждого человека мелатониновая кривая на удивление стабильна от ночи к ночи, а у разных людей одного пола и возраста кривые в деталях настолько отличаются, что можно говорить об индивидуальной кривой, характеризующей данную личность (рис.4).
Рис.4. Мелатониновые кривые у шести молодых здоровых испытуемых. По горизонтали - время суток в часах, по вертикали - концентрация мелатонина в плазме крови в пг/мл. Звездочками отмечено время отхода ко сну.Кроме суточных, существуют и сезонные ритмы колебания уровня мелатонина, причем не только у млекопитающих с сезонным циклом размножения, но и у человека. Посмертные исследования (аутопсия) показали, что у людей, живших в средних широтах Северного полушария и умерших в ноябре-январе, эпифизы достоверно больше по размеру и массе, чем у лиц, соответственно подобранных по возрасту, полу и месту проживания, умерших в мае-июле. Видимо, именно с ритмом эпифизарного мелатонина связаны в конечном счете сезонные изменения общей активности и эмоционального состояния человека (включая так называемые сезонные депрессии).Большая часть выброшенного в кровь гормона связывается с альбумином - основным белком плазмы. Таким способом мелатонин защищается от быстрого распада и транспортируется к клеткам-мишеням. По разным данным, период его полужизни в организме человека составляет от 30 до 50 мин. Свою активность мелатонин теряет в печени, где окисляется системой ферментов, связанных с белком Р-450, а затем выводится из организма.
Молекула основного эпифизарного гормона небольших размеров и высоко липофильна, в силу чего для нее не существует в организме никаких преград, включая плацентарный и гемато-энцефалический барьеры. С помощью меченного радиоактивными изотопами йода и трития мелатонина удалось обнаружить области его связывания во многих периферических органах и тканях млекопитающих: сетчатке, половых железах, селезенке, печени, вилочковой железе, желудочно-кишечном тракте, некоторых опухолевых тканях. Однако только в головном мозге выявлены особые многочисленные мембранные белки-рецепторы мелатонина, спаренные с гуанин-нуклеотид-связывающим белком (так называемым Г-белком). Здесь есть две главные области связывания: передняя часть гипофиза и супрахиазмальные ядра (СХЯ), причем у разных видов имеются заметные отличия. Так, у собак и кроликов рецепторы мелатонина присутствуют в обеих областях мозга, у копытных (крупного рогатого скота, лошади, осла) и куньих - только в гипофизе, а у человека - в основном в области СХЯ. Эти различия могут иметь важное значение как для сезонной репродукции, так и для часовых ритмов у дневных, ночных и сумеречных млекопитающих.
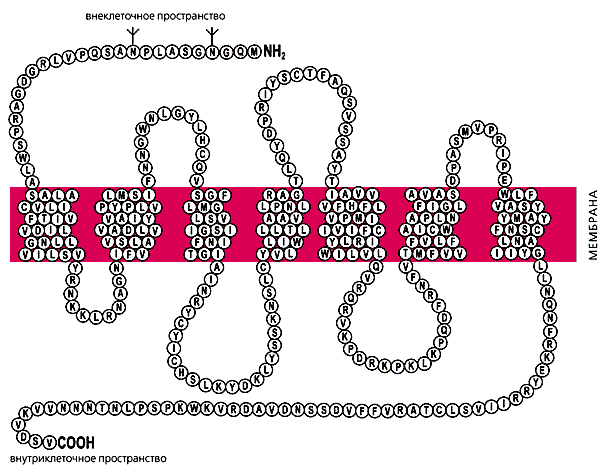
Недавно выделили ген, кодирующий основной белок-рецептор мелатонина, находящийся на мембране клеток-мишеней (рис.5). Взаимодействие гормона с рецептором угнетает активность этих клеток. Кроме этого, мелатонин может проникать сквозь мембрану, связываться с белками-рецепторами на поверхности ядра, даже проникать внутрь ядра и реализовывать свое действие на уровне ядерного хроматина, непосредственно влияя на синтез белка генетическим аппаратом клетки!
Рис.5. Структура белка-рецептора мелатонина. Каждый кружок представляет отдельный аминокислотный остаток, обозначенный общепринятым в биохимии однобуквенным кодом.Зародыши и новорожденные млекопитающие, включая человека, сами не образуют мелатонина, а пользуются материнским, поступающим через плаценту, а потом - с молоком матери. Секреция гормона начинается лишь на третьем месяце развития ребенка. С возрастом синтез мелатонина в эпифизе резко увеличивается и достигает максимума уже в первые годы жизни (не позднее 5 лет), а затем в течение всей жизни человека постепенно и плавно снижается (резкое падение наблюдается лишь в период полового созревания).Очевидно, что возрастная динамика мелатонина носит в основном адаптивный характер: ведь по мере ослабления выброса гормонов гипофизом и угасания деятельности периферических эндокринных желез потребность в их периодическом ночном торможении снижается и может вовсе исчезнуть. Недавно аутопсия подтвердила наличие в эпифизе рецепторов половых стероидов. Значит, эпифиз действительно получает обратную информацию о циркулирующих в крови гормонах. Сейчас это явление интенсивно изучается в ряде лабораторий мира.
У млекопитающих (включая человека) выброс эпифизарного мелатонина полностью контролируется супрахиазмальными ядрами. Эти небольшие парные образования и эпифиз млекопитающих - две половины главных биологических часов в нашем организме, находящиеся между собой во взаимотормозящих отношениях. Яркий свет стимулирует нейроны СХЯ, но тормозит выработку мелатонина эпифизом. В свою очередь, мелатонин, из-за высокой насыщенности высокочувствительными рецепторами СХЯ и смежных участков преоптической области, способен блокировать активность СХЯ - главного “генератора тактовых импульсов” в организме млекопитающих. При этом эпифизарный гормон взаимодействует с веществами, модулирующими активность супрахиазмальных ядер: нейромедиаторами (глутамат и серотонин) и нейропептидами (нейропептидтирозин и вещество П). Таким способом в системе внутрисуточной ритмики млекопитающих и человека поддерживается динамический гомеостаз.
Интересны идеи и разработки отечественного пинеолога (специалиста по эпифизу) А.М.Хелимского [5]. Он впервые указал, что социальный стресс (результат все ускоряющихся темпов и ритмов развития человеческого общества) стал главной движущей силой эволюции человека, которая реализуется через эпифиз и его основной гормон - мелатонин. По мнению Хелимского, хронический стресс матери во время беременности, столь характерный для больших городов, повышает уровень кортикостероидов (гормонов стресса), которые могут проникать через плаценту и подавлять у плода формирование эпифиза. За первую половину минувшего века средний вес эпифиза зрелого плода снизился, по его данным, почти в два раза! Такова, видимо, эпигенетическая (не связанная с наследственностью) реакция человеческой популяции на условия жизни в постиндустриальном обществе, с характерным действием стрессирующих факторов не только днем, но и ночью (залитые ярким светом ночные города - так называемый эффект Эдисона, ночной шум от автомобилей и самолетов, ночные будоражащие передачи по телевидению и пр.), и полным разрушением естественного для человека чередования периодов активности-покоя и сна-бодрствования.
Снижение угнетающего действия эпифизарного мелатонина на функции гипофиза усиливает выброс гормонов роста и стресса, половых гормонов, что проявляется в подростковой акселерации. В ней, быть может, и не было бы ничего плохого, если бы зачастую она не носила дисгармоничного характера. У подростков это выражается в диспропорциональном росте, раннем половом созревании, ожирении, гипертиреоидозах (патологическом разрастании щитовидной железы), усилении агрессивных реакций при фрустрации и т.п. Такова часть платы, которую вносит человечество за пренебрежение фундаментальными физиологическими факторами своего существования.
Мелатонин и сон
В настоящее время участие, по крайней мере косвенное, эпифизарного мелатонина в сезонной и внутрисуточной ритмике, сне-бодрствовании, репродуктивном поведении, терморегуляции, иммунных реакциях, внутриклеточных антиокислительных процессах, старении организма, опухолевом росте и психиатрических заболеваниях - не оставляет сомнений. Это доказано многочисленными исследованиями.
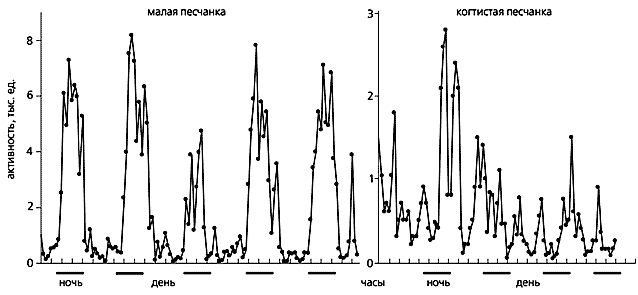
Рис.6. Суточные ритмы активности-покоя у двух родственных видов мелких грызунов при искусственном световом режиме: 12 часов света - 12 часов темноты. Слева - малая песчанка (ночная активность); справа - когтистая песчанка (сумеречная активность). По горизонтали - время (расстояние между двумя соседними штрихами соответствует 5 ч), по вертикали - двигательная активность в условных единицах. Горизонтальные штрихи под графиками - периоды выделения мелатонина (ночь).Однако не столь всестороннее до недавнего времени изучение влияния мелатонина на сон млекопитающих давало весьма противоречивые результаты. Известно, что звери по характеру своей активности подразделяются на дневных, ночных и сумеречных (не считая тех, чья активность не связана со сменой освещенности, например, крота-слепыша). У всех животных мелатонин выбрасывается эпифизом в темноте и блокируется на свету, а активность супрахиазмальных ядер подавляется мелатонином. Спрашивается, как может вещество, выделяющееся в одно и то же время, управлять столь непохожими типами поведения у разных видов млекопитающих (рис.6)? Пока окончательного ответа на этот вопрос нет, но очевидно, что мелатонин влияет на поведение косвенно, через какие-то еще не известные механизмы. В этой связи весьма важными представляются следующие недавно полученные данные:– межвидовые различия в распределении областей связывания мелатонина в головном мозге млекопитающих, о чём было сказано выше;Мы изучали действие небольших (физиологических) доз мелатонина на кроликах, животных с преимущественно сумеречной активностью. Исходя из литературных данных, за рабочую гипотезу приняли, что введение мелатонина в противофазе с суточным ритмом его продукции (т.е. в светлое время суток) должно вызывать обращение суточного хода кривой активности-покоя. Иными словами, ожидалось, что в нашей модели мелатонин лишь немного увеличит время бодрствования и уменьшит медленную и парадоксальную фазу сна.– различия в распределении подтипов рецепторов мелатонина внутри областей связывания;
– особая роль нейронов в области переднего гипоталамуса, находящейся в непосредственной близости к СХЯ и образующей с ними единый функциональный комплекс. В этой области обнаружены клетки, которые связаны с реальным ритмом активности-покоя у данного животного. Видимо, они преобразуют периодическую активность нейронов СХЯ, адаптируя ее к наиболее адекватному поведенческому ритму.
Однако на самом деле все оказалось гораздо сложнее: в некоторых сериях опытов мелатонин, действительно, подавлял сон, но в других - не влиял на него или даже увеличивал долю парадоксального сна (рис.7). По некоторым данным, в ряде случаев он также увеличивал парадоксальный сон у крыс, животных с выраженной ночной активностью. Аналогичное действие обнаружили и у некоторых здоровых испытуемых, когда после приема мелатонина скорее наступал быстрый (парадоксальный) сон и увеличивалась его доля в первых двух ночных циклах. Некоторые пациенты, которым вводили мелатонин по показаниям, отмечали появление необычайно ярких и эмоциональных сновидений.
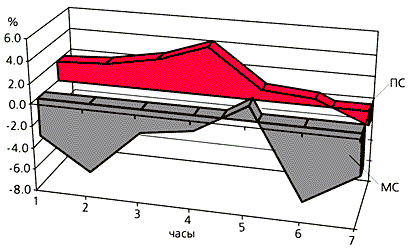
Рис.7. Эффекты малых доз мелатонина на сон кроликов: 50 мкг мелатонина, растворенного в 5 мл растительного масла, вводили через зонд в желудок кроликам через 3 ч после включения света в камере; в контроле тем же кроликам вводили растворитель. На основании 7-часовой непрерывной компьютерной записи полиграммы (совокупности физиологических показателей) определяли состояние животного в цикле сон-бодрствование. По горизонтали - время от момента введения в часах. По вертикали - различие между опытом и контролем в процентах (100% = 60 мин). По наклонной оси - прирост доли медленного сна (МС, отрицательный) и парадоксального сна (ПС, положительный - показано цветом).Причина разнообразного влияния мелатонина на сон не вполне ясна. Можно предположить, что его эффекты возникают вследствие изменения гормонального баланса и отражают взаимодействие с некоторыми важнейшими регуляторными пептидами, такими как соматолиберин, соматостатин, вазоинтестинальный полипептид и кортикотропиноподобный пептид из промежуточной доли гипофиза (CLIP).В то же время у диурнальных (дневных) млекопитающих, к которым относится человек, выброс мелатонина действительно совпадает с привычными часами сна. Это делает весьма привлекательной гипотезу о наличии и причинно-следственной связи между этими явлениями. У человека, однако, подъем уровня мелатонина не служит обязательным сигналом к началу сна. В различных лабораториях мира, в том числе и в исследованиях сотрудников Сомнологического центра Московской медицинской академии им. И.М.Сеченова, выполненных под руководством недавно ушедшего из жизни академика РАМН А.М.Вейна, прием мелатонина (от 0.3 до 3 мг) у большинства испытуемых вызывал лишь мягкий седативный эффект: способствовал некоторому общему расслаблению, снижал реактивность на обычные окружающие стимулы, что приводило к спокойному бодрствованию и плавному засыпанию. В отличие от сильных снотворных (феназепама, элениума, ивадала, имована и пр.), воздействующих на белки-рецепторы гамма-аминомасляной кислоты в мозге, мелатонин не вызывает ощущения невыносимой усталости и непреодолимой тяги ко сну. При необходимости человек легко преодолевает снотворные свойства мелатонина. Объективные и субъективные характеристики классических снотворных и мелатонина резко отличаются друг от друга.
Исходя из корреляции между субъективно ощущаемым и объективно подтвержденным ежевечерним нарастанием сонливости, с одной стороны, и увеличением концентрации мелатонина в крови, с другой, можно предположить, что он не прямо воздействует на сомногенные структуры головного мозга, а, скорее, создает некоторую “предрасположенность ко сну”, тормозит механизмы бодрствования. Благодаря высокой насыщенности СХЯ и смежных участков преоптической области высокочувствительными рецепторами, мелатонин, наряду с другими физическими (ярким светом) и вышеперечисленными биохимическими факторами, оказывает мощные модулирующие воздействия на активность главного осциллятора в организме млекопитающих, в том числе и человека. Так, при утреннем приеме он вызывает задержку фазы суточного ритма человека, а при вечернем - наоборот, сдвиг фазы вперед, причем не более чем на 30-60 мин/сут. Значит, ежедневно принимая мелатонин, можно сместить суточный цикл активности-покоя на несколько часов в ту или другую сторону. Такая потребность обычно возникает при трансмеридиональных авиаперелетах или сменной работе, когда сон нарушается и количественно, и качественно.
В отношении трансмеридиональных перелетов пинеологи дают следующие рекомендации:
– на расстоянии менее трех часовых поясов применение мелатонина бесполезно;Использование мелатонина для коррекции биоритмов при сменной работе зависит от ее характера, освещенности и особенностей данного субъекта. Вопросы о необходимости приема гормона, его дозы и времени решаются в каждом случае индивидуально с обязательным учетом формы кривой мелатонина (до и после применения). Разработанные в настоящее время весьма чувствительные (от 0.5 пг/мл) методы определения этого эпифизарного гормона не только в плазме крови, но и в моче и слюне, делают его использование вполне возможным.– при пересечении 3-6 поясов в восточном направлении принимать 0.2 мг мелатонина при отходе ко сну по местному времени для сдвига фазы суточного ритма “вперед”;
– при таком же расстоянии в западном направлении принимать 0.1 мг мелатонина сразу после полуночи по местному времени, если человек в это время не спит, или же при спонтанном пробуждении в ранние утренние часы, характерном при таких перелетах, для сдвига суточной фазы “назад”;
– при пересечении 7-12 поясов в любом направлении мелатонин противопоказан, поскольку может усилить дизритмию - так называемый внутренний десинхроноз - и ухудшить субъективное состояние человека.
Несколько лет назад появились многочисленные публикации о “безвредности” и даже необходимости постоянного возмещения возрастной “нехватки” мелатонина. Это якобы должно улучшить общее состояние здоровья пожилых людей и продлить жизнь [6]. Такое совершенно необоснованное с позиций современных знаний представление привело к беспрецедентному в истории фармацевтики явлению: в США гормон человека - синтетический мелатонин - был рекомендован в качестве пищевой добавки. Бесконтрольное по сути использование препарата, повышающее его концентрацию в десятки, сотни и даже тысячи раз по сравнению с естественным ночным уровнем, может не только нарушить суточный ритм и цикл сон-бодрствование, но и вызвать общую эндокринную недостаточность из-за неадекватного и чрезмерного торможения гормонов гипофиза и периферических эндокринных желез тогда, когда в подобном торможении уже нет нужды. Говоря другими словами, чрезмерная еженощная концентрация мелатонина может ухудшить здоровье и укоротить жизнь - т.е. привести к результатам, прямо противоположным тем, которые декларируются адептами “мелатонинового чуда”!
Мелатонин абсолютно не токсичен, что способствовало его применению в очень больших, так называемых фармакологических дозах (от миллиграммов до граммов), далеких от физиологических (они составляют, как уже говорилось, десятые доли миллиграмма). Некоторые фирмы, поставляющие синтетический мелатонин на фармацевтический рынок, в том числе на российский, перешли на расфасовку мелатонина исключительно в дозах 3 и 5 мг активного вещества на таблетку или капсулу. Однако только прием физиологических доз препарата (0.1-0.3 мг), обеспечивающих подъем мелатонина в плазме крови до 50-120 пг/мл (средний ночной уровень у взрослых), вызывает мягкий снотворный эффект. Во многих случаях у людей, страдающих бессонницей, малые дозы мелатонина улучшают ночной сон, не изменяя его структуры. Это говорит о том, что больничным и аптечным учреждениям необходимо отказаться от закупок коммерческого мелатонина и использовать препарат в расфасовке по 0.1-0.3 мг.
Итак, несмотря на огромное количество работ, посвященных этому вопросу, не будет ошибкой сказать, что эпифиз остается наименее изученной из всех эндокринных желез, а фундаментальная роль мелатонина только начинает осознаваться [1-5]. Это вещество, с помощью которого жизнь на Земле, возникшая благодаря Солнцу, защищалась от его губительного воздействия, оказалась важнейшим “тормозным” элементом в гормональной системе млекопитающих.
Литература
1. Анисимов В.Н., Кветной И.М., Комаров Ф.И., Малиновская Н.К., Рапопорт С.И. Мелатонин в физиологии и патологии желудочно-кишечного тракта. М., 2000.
2. Мелатонин в норме и патологии / Под ред. акад. РАМН Ф.И.Комарова. М., 2003 (в печати).
3. Слепушкин В.Д., Пашинский В.Г. Эпифиз и адаптация организма. Томск, 1982.
4. Чазов Е.И., Исаченков В.А. Эпифиз: место и роль в системе нейроэндокринной регуляции. М., 1974.
5. Хелимский А.М. Эпифиз. М., 1969.
6. Пьерпаоли В., Регельсон У. Чудо мелатонина / Пер. с англ. М., 1997.